Article Text

Abstract
A multitude of physiological effects and putative pathophysiological roles have been proposed for the endogenous cannabinoid system in the gastrointestinal tract, liver and pancreas. These range from effects on epithelial growth and regeneration, immune function, motor function, appetite control, fibrogenesis and secretion. Cannabinoids have the potential for therapeutic application in gut and liver diseases. Two exciting therapeutic applications in the area of reversing hepatic fibrosis as well as antineoplastic effects may have a significant impact in these diseases. This review critically appraises the experimental and clinical evidence supporting the clinical application of cannabinoid receptor-based drugs in gastrointestinal, liver and pancreatic diseases. Application of modern pharmacological principles will most probably expand the selective modulation of the cannabinoid system peripherally in humans. We anticipate that, in addition to the approval in several countries of the CB1 antagonist, rimonabant, for the treatment of obesity and associated metabolic dysfunctions, other cannabinoid modulators are likely to have an impact on human disease in the future, including hepatic fibrosis and neoplasia.
Statistics from Altmetric.com
The marijuana plant Cannabis has been used to treat symptoms and diseases including anorexia, emesis, abdominal pain, gastroenteritis, diarrhoea, intestinal inflammation and diabetic gastroparesis.1–6 Δ9-Tetrahydrocannabinol (Δ9-THC), the main active ingredient of Cannabis, activates two Gi/o-coupled membrane receptors, named CB1 and CB2 receptors. Cannabinoid CB1 receptors are located primarily on central and peripheral neurons. In the gut, CB1 receptors modulate neurotransmitter release, whereas CB2 receptors are mainly associated with immune functions.7 CB2 receptors are also expressed in brain areas implicated in emesis.8
Endocannabinoids are endogenous ligands that activate cannabinoid receptors in mammalian tissues. The most extensively studied are anandamide and 2-arachydonylglycerol (2-AG). These are lipid mediators synthesised “on demand” from membrane phospholipids, released from cells immediately after production, activate receptors to elicit a biological response, after which they are inactivated through reuptake (facilitated by the putative endocannabinoid membrane transporter (EMT)) and enzymatic degradation. Anandamide is inactivated by fatty acid amide hydrolase (FAAH),9–12 and 2-AG by monoacylglycerol lipase (MAGL) (see fig 1 for further details). Additionally, FAAH may also metabolise 2-AG.8

Apart from effects on CB receptors, endocannabinoids also activate the transient receptor potential vanilloid type 1 (TRPV1), which is mainly expressed by primary afferent neurons and the orphan G protein-coupled receptor GPR55.13–15 Intestinal endocannabinoids activate the CB1 receptor (in physiological and pathophysiological states), the CB2 receptor (in pathophysiological states only) and the TRPV1 receptor (in some inflammatory conditions such as bacterial enteritis).16
Given the multitude of physiological effects and putative pathophysiological roles proposed for the endogenous cannabinoid system in the gut, cannabinoids have the potential to be a panacea for gut and liver diseases. This review critically appraises the experimental and clinical evidence supporting application of cannabinoid-based drugs in gastrointestinal, liver and pancreatic diseases.
THE ENDOGENOUS CANNABINOID SYSTEM IN THE GUT
The endogenous cannabinoid system has been detected in the gut, including cannabinoid receptors, endocannabinoids and proteins involved in endocannabinoid biosynthesis (eg, N-acyl-phosphatidylethanolamine-selective phospholipase D (NAPE-PLD) see also fig 1) and inactivation (eg, FAAH).
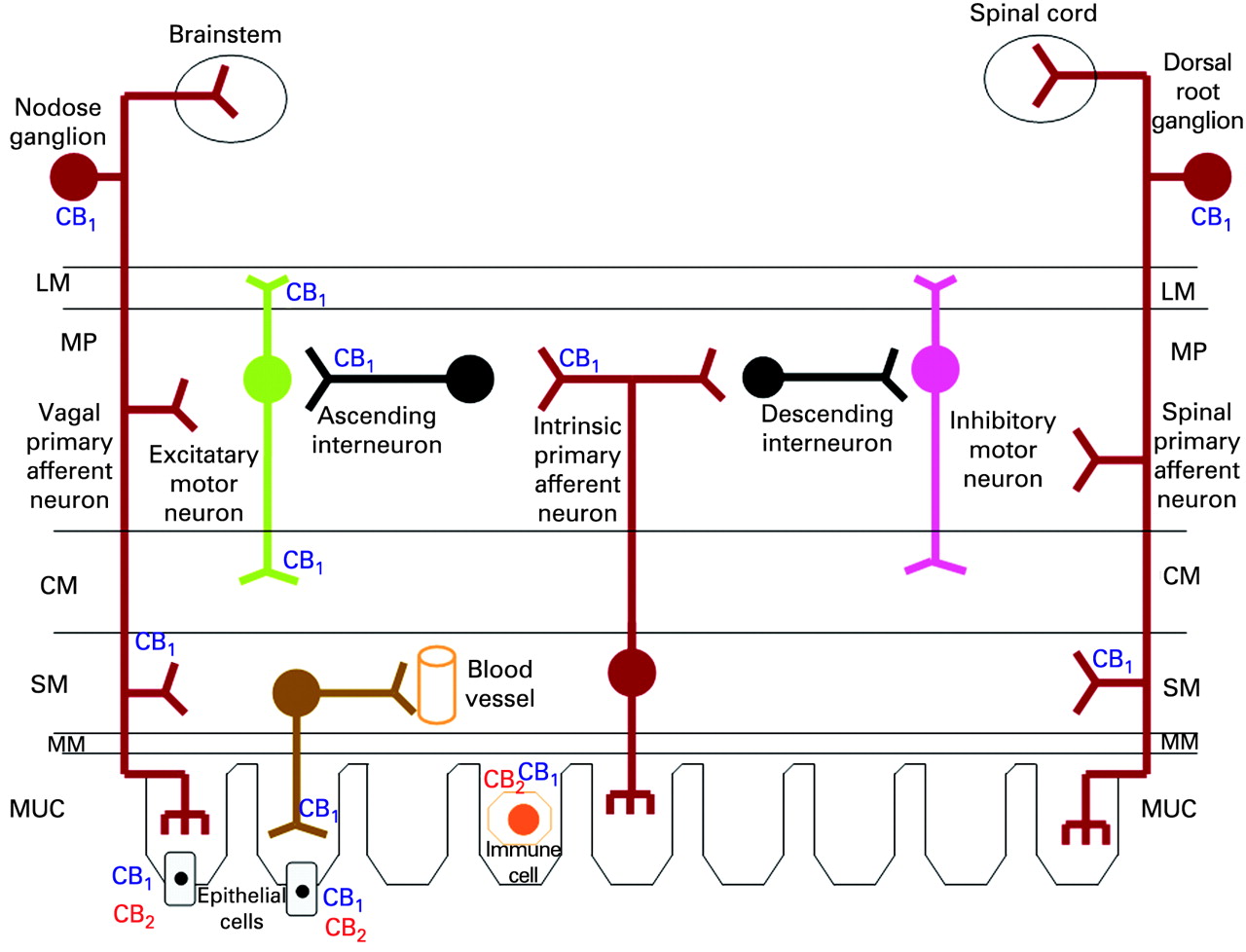
Localisation of cannabinoid receptors in the gut
CB1 receptors are located throughout the gastrointestinal tract,17 mainly in intrinsic (enteric) and extrinsic neurons1 such as the cell bodies of sensory neurons in the dorsal root ganglia and nodose ganglion, and vagal efferents.18–28 The enteric nervous system (fig 2) is the major site of action of cannabinoids in the digestive tract.1 2 In animal and human intestine, CB1 receptors have been identified on cholinergic nerves in the myenteric and submucosal plexus, in nerve fibres innervating the circular and longitudinal muscles,1 29 30 in non-cholinergic neurons containing substance P or vasoactive intestinal peptide, and in non-neuronal cells, including crypt epithelial cells and smooth muscle cells.1 2 29 30

CB2 receptors are expressed by lamina propria plasma cells and activated macrophages, possibly by epithelial cells29 and by myenteric and submucosal plexus ganglia in the normal human ileum.31 The role of neuronal CB2 receptors awaits further investigation.
CB receptors in disease
CB1 and CB2 receptors may be upregulated (table 1) in experimental models of inflammatory bowel disease and diarrhoea,25 26 32–34 and in human diseases.29 35–39 In untreated coeliac disease, CB1 receptors are upregulated in the subepithelial region, where gluten-reactive proinflammatory Th1 cells are located.35 Intense epithelial CB1 and CB2 immunoreactivity was observed at sites of mucosal ulceration in inflammatory bowel diseases.29 35
Endogenous ligands
Levels of 2-AG are ∼200-fold higher than those of anandamide in human small and large intestine;37 38 the basal levels of both endocannabinoids in the human gut are so high (in the μM range for 2-AG) that they would permanently activate cannabinoid receptors2–39 if they were not efficiently inactivated. Intestinal endocannabinoid (mainly anandamide) levels increase after noxious stimuli in animals35 37 40 41 and in intestinal biopsies of patients with colon cancer,36 diverticulitis38 and coeliac disease35 (table 1). Anandamide levels normalise with remission of coeliac disease, suggesting that intestinal anandamide may protect against inflammation. Endocannabinoids are reduced in animals after administration of the emetic drug cisplatin42; it has been suggested that endocannabinoid reduction may contribute to the emesis associated with cisplatin.39
The endogenous source and the biosynthetic pathways for endocannabinoid production have been poorly investigated in the gut. Endocannabinoids are produced by both the serosal and mucosal side of the human intestine,37 and studies on cell lines have shown that human intestinal epithelial cells can produce endocannabinoids36; additional sources of endocannabinoids in the gut include human endothelial cells,43 platelets,44 resident macrophages and other inflammatory/immune cells1 such as lymphocytes.45 Although sensory nerve stimulation can release anandamide in cultured dorsal root ganglion neurons,46 there is no direct evidence to date that enteric nerves synthesise endocannabinoids.
Proteins involved in endocannabinoid biosynthesis and inactivation
The enzyme for anandamide synthesis (NAPE-PLD) is found in rodent stomach antrum and fundus,47 enterocytes and lamina propria cells of the small intestine.48 FAAH (degradation enzyme) is expressed in different regions of the rodent gut,32 33 49 50 and increased during experimental inflammation.32 33 Functional studies suggest that EMT might be involved in experimental inflammation,37 ileus25 and diarrhoea.26
PHARMACOLOGICAL ACTIONS OF ENDOCANNABINOIDS IN THE GASTROINTESTINAL TRACT
Endocannabinoids exert multiple pharmacological actions in the gut, chiefly by CB1 and/or CB2 receptor activation, and, in addition, TRPV1 or non-receptor mechanisms.
Effects on CB receptors
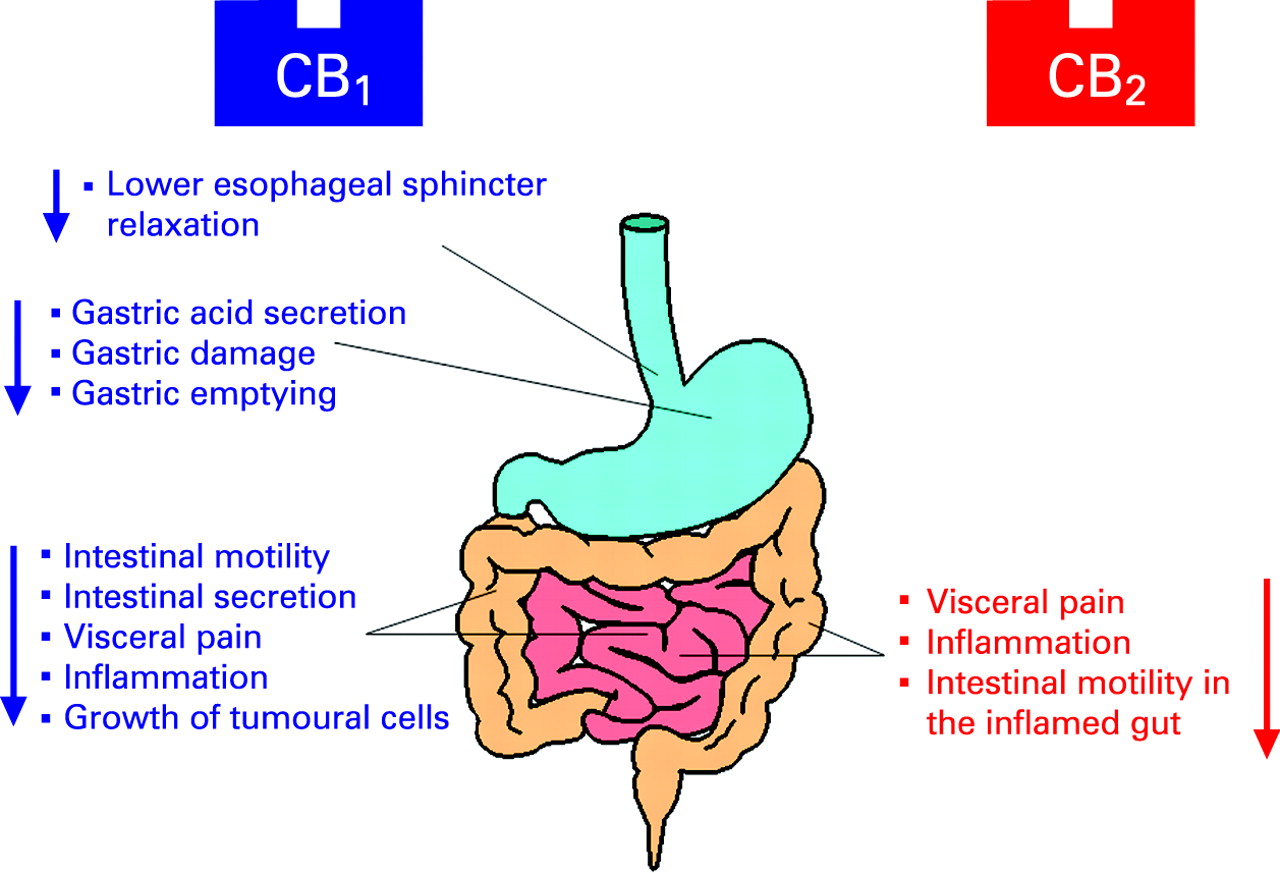
In vivo pharmacological studies (fig 3) in rodents show that CB1 receptor agonists increase food intake,51 52 but inhibit emesis,53–56 gastric acid secretion.23 57 experimentally induced gastric ulcers,58–60 relaxation of the lower oesophageal sphincter (LOS),61 62 gastric emptying,63 64 intestinal motility27 65 and secretion,26 exert antineoplastic actions,41 attenuate visceral pain66 and reduce inflammation.33 34 37 CB2 receptor activation may also affect visceral pain and inflammation.34 66 67

Conversely, CB1 blockade may be emetic,68 reduce food intake,51 52 increase gastric emptying69 and intestinal motility,27 65 and exacerbate inflammation,33 visceral pain66 and diarrhoea.26
Cannabinoid receptors may also be indirectly activated by inhibitors of endocannabinoid inactivation or reuptake, which increase levels of endocannabinoids and result in anti-inflammatory37 and antidiarrhoeal effects.26 Conversely, FAAH inhibition elicits anticancer41 and anti-inflammatory effects33 37 and reduces gastrointestinal motility.50 69
Effects on non-cannabinoid receptors
TRPV1 expressed by primary afferent neurons70 71 is activated by high concentrations of anandamide, resulting in enteritis40 in the rat in vivo and enhanced acetylcholine release from myenteric guinea pig nerves.72 TRPV1 activation in the dorsal motor nucleus of the vagus and area postrema has antiemetic effects.56
Finally, anandamide may inhibit acetylcholine-induced myogenic contractions in the human intestine independent of cannabinoid and TRPV1 receptors.73
POTENTIAL THERAPEUTIC APPLICATIONS IN THE GUT
The potential therapeutic applications of cannabinoids are based on experimental and clinical studies.
Anorexia and obesity
Endogenous cannabinoids may regulate energy balance and food intake by central effects in the limbic system (which is involved in evaluation of pleasure associated with foods) and the hypothalamus (stimulation of food intake), and by peripheral effects, including the adipose tissue, skeletal muscles, pancreas, liver and the gastrointestinal tract.74 75 Plasma endocannabinoids are increased in overweight/obese patients and in underweight women with restricting anorexia nervosa, but not in patients with binge-eating disorders.76–79 Furthermore, restricting anorexia nervosa and binging/purging anorexia nervosa may be associated with different alleles of the CB1 receptor gene (CNR1).80 In several species including humans, CB1 agonists increase food intake and promote body weight gain, whereas selective CB1 receptor antagonists may reduce food intake and body weight.51 52
Food deprivation increased anandamide (but not 2-AG) levels and vagal CB1 receptor expression in the rat small intestine; these changes were normalised by re-feeding. CB1 receptors on afferent sensory neurons may be involved in modulation of appetite; and anandamide acts as a “hunger signal” in the intestine.19 24 81 82
Two systematic reviews and meta-analyses83 84 analysed four high-quality, 12- to 24-month, multicentre randomised placebo-controlled trials85–88 of the CB1 receptor antagonist rimonabant for the treatment of obesity and metabolic disorders in a total of 4105 individuals. Rimonabant therapy (20 mg/day), in association with a low-caloric diet, resulted in a greater reduction in body weight of 4.7 kg (average) over placebo. There are reports of a twofold increase in the incidence of psychiatric disorders (depression, anxiety, irritability, aggression); thus physicians need to be aware of this risk and to be alert to the development of such psychiatric disorders in patients receiving rimonabant.83 84 Adverse effects associated with rimonabant include nausea and diarrhoea, consistent with the pharmacological actions of rimonabant in animals on gut functions (prokinetic and emetic).
Cannabinoids stimulate appetite in human volunteers89 and AIDS patients,90 but not in patients with advanced cancer.91
Nausea and vomiting
Cannabinoids prevented experimentally induced emesis92 by activation of vagal CB1 receptors related to gastric function either centrally, in the area postrema and dorsal vagal complex, or peripherally on abdominal vagal efferents. Activation of brainstem TRPV156 and CB2 receptors7 93 94 may also have antiemetic effects. Pharmacological studies revealed proemetic effects of CB1 receptor antagonists68 92 and antiemetic actions of endocannabinoid re-uptake42 and FAAH inhibitors,95 96 suggesting that these receptors may be physiologically involved in the genesis of nausea and vomiting.
The synthetic cannabinoid, nabilone, and dronabinol (Δ9-THC) are available for prescription as antiemetics for co-administration with cancer chemotherapeutic agents and are slightly superior to conventional antiemetics (such as prochlorperazine or metoclopramide, or 5-HT3 antagonists) for chemotherapy-induced emesis.97 98 Side effects are common with cannabinoids. Some may be beneficial (euphoria, “high”, sedation); others are harmful (dysphoria, depression, hallucinations). Many patients have a strong preference for cannabinoids.98
A clinical syndrome called cannabinoid hyperemesis is characteristically associated with cyclical vomiting with use of cannabis, and relief of symptoms in hot showers.99 100 Clinical experience (unpublished by one of the authors) suggests that symptoms may be exacerbated by withdrawal from cannabis—for example, when college or university students are visiting their families during holidays. The mechanism causing this syndrome and leading to relief with compulsive bathing, are unclear. The non-selective cannabinoid agonist, dronabinol, results in modest retardation of gastric emptying of solids101 102; this is unlikely to explain the hyperemesis.
Gastrointestinal reflux disease
Two mechanisms involved in gastro-oesophageal reflux (GOR) disease are transient LOS relaxations and gastric acid production; both may be influenced by cannabinoid drugs. CB1 agonists inhibited LOS relaxation and GOR in dogs,61 and LOS relaxation in ferrets,62 through central and peripheral vagal mechanisms.61
Cannabinoids decrease acid production in rodents through activation of CB1 receptors on vagal efferents to gastric mucosa,23 57 not by direct effects on parietal cells. CB1 receptor activation is protective in animal models of gastric ulcers.58–60
Irritable bowel syndrome (IBS)
There is a significant association of C825T variation in the FAAH gene with symptom phenotype in diarrhoea-predominant IBS (D-IBS) and the mixed bowel function IBS phenotypes, and with faster colonic transit in D-IBS.103 These data support the hypothesis that cannabinoid mechanisms may play a role in the control of colonic motility in humans (discussed further below).
Cannabinoids may be potentially beneficial for IBS patients by inhibition of intestinal motility, secretion and visceral sensation, mediated, at least in part, by activation of enteric CB1 receptors. CB2 receptor stimulation may reduce visceral pain. However, given the complexity of the pathophysiology of IBS, and the incomplete predictability of animal models, evidence for clinical application of cannabinoid drugs in IBS is still at an early stage; it is important to note there is potential for associated central unwanted effects (see later).
Intestinal motility
Cannabinoid agonists act on prejunctional CB1 receptors to reduce excitatory (eg, cholinergic) enteric transmission in different regions of the gut, including the human intestine.30 38 104 Other mechanisms may also be involved—for example, inhibition of non-cholinergic excitatory transmission in the guinea pig ileum105 106 and mouse colon,107 and reduction of inhibitory (non-nitrergic) transmission in the mouse colon.28 CB1 receptor activation constitutes a physiological “brake” for the small and large intestine. This is based on several lines of evidence25 27 32 50 108 109 (summarised in the box beolw). Cannabinoids also reduce motility in the inflamed gut, possibly mediated by CB2 receptors110 and myenteric CB1 receptors whose expression is increased by inflammation.32 111
Evidence supporting important actions of cannabinoids on motility, inflammation and fibrogenesis
PHYSIOLOGICAL BRAKE of the rodent small and large intestine
High concentrations of endocannabinoids in intestinal tissues32
CB1 receptor agonists inhibit intestinal motility (not via CB2 or TPRV1)108
CB1 receptor antagonist rimonabant stimulates intestinal motility27 32
Increased transit is observed in CB1-deficient mice109
INTESTINAL ANTI-INFLAMMATORY ACTION IN VIVO
Activation of both CB1 and CB2 receptors reduced colon weight gain, colon shrinkage, colon inflammatory damage score and diarrhoea induced by experimental colitis34
Genetic ablation of CB1 receptors or CB1 receptor antagonists rendered mice more sensitive to colitis33
FAAH-deficient mice as well as inhibitors of anandamide reuptake or enzymatic hydrolysis (all with presumed higher levels of anandamide) showed protection against colonic inflammation33 35
Anandamide levels and expression of CB receptors are increased in the inflamed colon33 35
CB1 and CB2 receptor activation reduces hypersensitivity and pain induced by experimental colitis66
HEPATIC FIBROGENIC AND VASCULAR EFFECTS
Monocytes of rats with cirrhotic ascites are activated to produce anandamide which contributes to arterial hypotension127 137
Circulating anandamide is elevated in cirrhotic animals and humans127 132 137
Endocannabinoids are increased in the cirrhotic liver136
CB1 and TRPV1 receptors are upregulated in hepatic vascular endothelial cells of cirrhotic rats127 128 137
Blockade of CB1 and TRPV1 receptors counteracted systemic hypotension and elevated mesenteric blood flow and portal pressure in rats with experimental cirrhosis127 128 137
Anandamide (via CB1 receptor and TPVR1 activation) increased mesenteric flow and vasodilated mesenteric arteries in cirrhotic rats with ascites,126 150 and increased (via thromboxane A2 production) the intrahepatic resistance in cirrhotic rats150 151
In healthy human volunteers, cannabinoid agonists reduce gastric emptying112 and increase colonic compliance and reduce colonic motility, particularly phasic and tonic motor responses to meal ingestion101 (fig 4).

Intestinal secretion
In Ussing chamber studies of muscle-stripped intestine, activation of CB1 receptors on submucosal neurons and extrinsic primary afferents in the submucosa reduces ion transport.21 113 114 In vivo, CB1 receptor activation inhibits cholera toxin-induced secretion in mice26 and castor oil-induced diarrhoea in rats.115 This is consistent with the traditional use of Cannabis in the treatment of cholera and dysentery.116
Visceral sensation and pain in the normal and in the inflamed gut
There is strong evidence that cannabinoids have the potential to relieve pain, with most experimental data related to somatic pain.6 However, the role of cannabinoids in the control of visceral nociception only recently has been explored. CB1 and CB2 receptor activation inhibits abdominal sensitivity to colorectal distension (a surrogate of visceral pain) in rats under basal conditions.66 This contrasts with the increased colonic sensation to distension observed in humans exposed to relatively low doses of the non-selective CB agonist, dronabinol.101
In the inflamed gut, activation of either CB1 or CB2 receptors causes reduction of hyperalgesia at doses lower than those active in the normal gut.66 CB2 receptor activation also reduces the visceral response due to the algesic compound bradykinin.117 This is particularly relevant since peripheral activation of primary afferent fibres by algesic chemicals released in the gut is frequently the cause of visceral pain.118 Additionally, CB1 and CB2 receptor activation was antinociceptive in a mouse model of acute pancreatitis, supporting the potential of cannabinoids in limiting visceral pain.119
The role of the endocannabinoid system in endogenous antinociceptive pathways has been investigated with the CB1 receptor antagonist, rimonabant, which enhanced colitis-induced hyperalgesia.66 It is worth noting that this effect was not observed with a CB2 receptor antagonist or in control animals.66 These results suggest involvement of the endocannabinoid system in inflammatory hyperalgesia, through CB1 receptors.
In a rodent model, epithelial CB2 receptors were upregulated by a probiotic, Lactobacillus acidophilus,120 and this is associated with increased sensation threshold (decreased pain) to elicit colorectal distension-induced visceromotor responses. This may be relevant to the potential therapeutic benefit of probiotics in IBS.121
In summary, the potential of CB modulation of visceral pain in humans, such as those with IBS which at most has a minor inflammatory component, is unclear. From animal studies, CB1 mechanisms may be most consistent, though data from different models suggest that CB2 may also be worthy of further study. The effects on colonic motility in humans and the observation of diarrhoea as a side effect in the large rimonabant trials suggest that the effect of cannabinoids on colonic motility may be more relevant in IBS.
Inflammatory bowel disease
Patients with inflammatory bowel disease (IBD) anecdotally report symptom relief from smoking marijuana. There are increased endocannabinoid levels and CB1 and CB2 receptor expression in biopsies from patients with IBD (table 1). In well-established models of IBD in rodents (ie, colitis induced by dextran sulfate, dinitrobenzene sulfonic acid (DNBS) or mustard oil), several lines of evidence suggest endocannabinoids may limit intestinal inflammation in vivo via CB1 and/or CB2 receptor activation (see box).33–35
CB1 and CB2 receptors modulate inflammatory processes. CB1 receptors promote epithelial wound healing,29 and CB2 receptor activation inhibits interleukin 8 release in human colonic epithelial cells.122
Cannabinoid agonists may also reduce motility in the inflamed intestine; both CB1 (eg, in ileitis induced in mice by the irritant croton oil) and CB2 receptors may be involved.32 110 CB1-deficient mice lack early protective mechanisms against neuromuscular changes initiated in the distal colon by DNBS administration.111 In contrast, in lipopolysaccharide-induced intestinal hypermotility in the rat, hypermotility was normalised by a CB2 agonist, but not by a CB1 receptor agonist.110
In summary, cannabinoids reduce response to gut inflammation via CB1 and/or CB2 activation by direct suppression of proinflammatory mediators, inhibition of intestinal motility and diarrhoea, and attenuation of visceral sensitivity (fig 5).

THE ENDOGENOUS CANNABINOID SYSTEM IN THE LIVER
Cannabinoid receptors
CB1 and CB2 receptors (which are respectively faintly or not expressed in normal livers) are substantially upregulated in experimental liver injury and in liver cirrhosis of various aetiologies123–135 (table 2). Upregulation occurs in non-parenchymal cells distributed along the fibrous septa in spindle-shaped cells, inflammatory cells and ductular proliferating cells. CB2 receptors are also identified in inflammatory cells and bile duct epithelial cells along fibrotic septa,133 and in hepatocytes of non-alcoholic steatosis and steatohepatitis.130 CB1 and CB2 receptors are expressed in activated (but not non-activated) hepatic stellate cells (the main fibrogenic cells in the liver) and in myofibroblasts in human liver.136
Endocannabinoids and regulating enzymes
Both anandamide and 2-AG have been identified in the liver, with higher levels in animal models of liver damage.135 136 Plasma (or circulating monocyte) anandamide levels are increased in patients with liver cirrhosis133 137 138 (table 2).
Experimentally, endotoxin-induced anandamide synthesis in macrophages has been implicated in the hypotension of septic shock and advanced liver cirrhosis.9
FAAH is highly expressed in primary hepatocytes, which were completely resistant to anandamide-induced cell death, whereas primary hepatic stellate cells expressed low levels of FAAH and were highly sensitive to anandamide-induced cell death.139
PATHOPHYSIOLOGY AND POTENTIAL THERAPEUTIC APPLICATIONS OF THE CANNABINOIDS IN LIVER DISEASES
Endocannabinoids appear to be involved in the pathophysiology of acute and chronic liver disease, including inflammation, fibrogenesis, steatosis, vascular complications and neurological dysfunction associated with liver diseases (evidence summarised in the box).140–144 Potential therapeutic applications of cannabinoid drugs in liver diseases are illustrated in table 3 and fig 6.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Fibrosis-associated chronic liver diseases
Endocannabinoids may impact liver fibrogenesis via cannabinoid receptor-dependent and independent mechanisms. Cannabinoid CB1 and CB2 receptor mechanisms induce profibrogenic and antifibrogenic effects, respectively, suggesting that combining CB2 agonists and CB1 antagonists holds promise for treatment of fibrosis in chronic liver diseases.
Profibrogenic CB1-mediated effects
Blockade of CB1 receptors decreased the wound healing response to acute liver injury and inhibited progression of fibrosis in three models of chronic liver injury. Reduced accumulation of hepatic myofibroblasts after growth inhibition and enhanced apoptosis were major determinants of the antifibrotic effect of CB1 receptor antagonism.131
Antifibrogenic CB2-mediated effects
CB2-deficient mice developed increased liver fibrosis following exposure to carbon tetrachloride, compared with wild-type mice.133 Moreover, pharmacological activation of CB2 receptors leads to reduction of hepatic collagen content in rats with pre-existing carbon tetrachloride-induced cirrhosis.125 The reduced collagen is associated with reduced density of monocyte/macrophages within and around the portal tracts, decreased numbers of α-smooth muscle actin-positive cells, increased apoptosis in myofibroblasts and monocytes, diminished tissue content of collagen I and α-smooth muscle actin and increased proteolytic enzyme, matrix metalloproteinase-2.125
In isolated hepatic myofibroblasts and activated hepatic stellate cells, activation of CB2 receptors elicited antifibrogenic properties by triggering growth inhibition and apoptosis. Growth inhibition involved cyclooxygenase-2 (COX-2), and apoptosis resulted from oxidative stress.133
Non-receptor-mediated effects of cannabinoids on fibrogenesis
Endocannabinoids may also modulate fibrogenesis in the liver through mechanisms that are independent of CB1, CB2 or TRPV1 receptors.136 145 Anandamide and 2-AG induced necrosis in hepatic stellate cells (which express low levels of FAAH), but not in hepatocytes, which have high expression of FAAH.139 Endocannabinoid-induced cell death involves membrane cholesterol, generation of reactive oxygen species and intracellular Ca2+. Liver concentrations of 2-AG, but not anandamide, increased in two experimental models of hepatic fibrogenesis (ie, carbon tetrachloride and bile duct ligation) and reached tissue levels of 2-AG that are consistent with those that induce cell death in vitro.136 140 These results suggests that 2-AG, rather than anandamide, is important in fibrogenesis and that drugs which selectively increase hepatic 2-AG may be pursued as a therapeutic strategy for hepatic fibrosis as they have anti-inflammatory actions and activate apoptosis rather than fibrosis.
Anandamide may also induce apoptosis in human hepatocytes undergoing rapid proliferation; it has been proposed that this endocannabinoid may counteract liver regeneration, which may be deleterious;146 conversely, it is conceivable that inhibition of anandamide or CB1 receptors may enhance liver regeneration.
Cannabis use as a risk factor for progression of fibrosis
Chronic use of marijuana was associated with hepatic morphological and enzymatic alterations.147 Two studies involving 270 and 204 patients found significant associations between daily cannabis smoking and progression of fibrosis148 or with development of moderate to severe fibrosis in chronic hepatitis virus C infection.149
Vascular and neurological changes associated with liver cirrhosis
Endocannabinoids have been implicated in haemodynamic and neurological consequences associated with liver cirrhosis, including portal hypertension, encephalopathy and cardiomyopathy (fig 6).
Portal hypertension
Upregulated CB1- (and possibly TRPV1) dependent cannabinoid mechanisms contribute to the pathogenesis of portal hypertension and associated systemic hypotension. This is supported by evidence126–128 132 137 150 151 summarised in the box.
Cirrhotic cardiomyopathy
In vitro evidence suggests that activation of cardiac CB1 receptors by endogenous anandamide, which is elevated in the hearts of cirrhotic animals, contributes to reduced cardiac contractility in liver cirrhosis.152 153 The blunted cardiac contractility in cirrhosis is restored toward normal by a CB1 antagonist.152
Hepatic encephalopathy
In an animal model of acute hepatic failure, neuroprotection may be achieved through CB1 blockade or CB2 activation.154 Cannabinoids ameliorate cerebral dysfunction via AMP-activated protein kinase.155
Hepatic ischaemia
In an in vivo model of hepatic ischaemia–reperfusion injury, endocannabinoids increased in the liver and in the plasma (table 2).135 156 Activation of CB2 receptors protects against hepatic damage by decreasing inflammatory cell infiltration and production of inflammatory mediators. In contrast, CB2-deficient mice developed increased ischaemia–reperfusion-induced tissue damage and a proinflammatory phenotype. These results suggest that oxidative and/or inflammatory stimuli may trigger endocannabinoid production, and that targeting CB2 receptors may protect against hepatic ischaemia–reperfusion injury.157 158
Steatosis and steatohepatitis
Blockade of CB1 receptors prevents development of steatosis in obese mice.159 The CB1 receptor antagonist, rimonabant, decreased hepatomegaly, completely reversed hepatic steatosis, decreased plasma circulating levels of enzyme markers of liver damage and reduced hepatic tumour necrosis factior α (TNFα) levels that characterise steatohepatitis.160 161 Daily cannabis smoking has been identified as an independent predictor of the severity of steatosis in patients (n = 315) with chronic hepatitis C.162 This clinical observation supports the experimental data160 161 suggesting that CB1 antagonists may be pursued as a novel therapy for non-alcoholic fatty liver disease (NAFLD).
Cholestatic pruritus
Δ9-THC (2.5 mg at bedtime) relieved intractable pruritus due to cholestasis and improved sleep in three patients refractory to conventional therapies.163 It has been hypothesised that efficacy of Δ9-THC in pruritus is due to its antinociceptive actions; experimentally, a synthetic cannabinoid increased the threshold to nociception in rats with cholestasis secondary to bile duct resection.164
Cholangiopathies
Cholangiopathies are diseases affecting the biliary system and they share a number of pathogenetic mechanisms, including inflammation, cholestasis, fibrosis, apoptosis, altered development and neoplastic transformation,165 which target intrahepatic bile duct epithelial cells (ie, cholangiocytes).166 Proliferation of cholangiocytes represents a key mechanism of repair responsible for maintaining the integrity of the biliary tree,167 and it occurs in virtually all pathological conditions of liver injury where it is associated with inflammation, regeneration and repair.167 In a mouse model of cholestatic liver disease, anandamide inhibited cholangiocyte proliferation via a CB2-dependent mechanism, an effect associated with increased accumulation of reactive oxygen species.168 These results suggest that modulation of the endogenous cannabinoid system may be proposed for the treatment of cholangiopathies
ACUTE PANCREATITIS
CB1 and CB2 receptor immunoreactivity was weakly revealed in pancreatic acini, nerves, blood vessels and duct cells in the pancreas of normal subjects.120 169 However, in specimens of subjects with acute pancreatitis, vital pancreatic parenchyma displayed a striking increase in immunostaining of CB1 in duct and a moderate increase of CB1 and anandamide (but not 2-AG) in nerves and acini as compared with normal pancreas; acinar and ductal CB2 receptors were increased to a lesser extent than CB1 receptors.120 Acinar cells in the vicinity of severe necrosis in human acute pancreatitis specimens revealed intense cytoplasmic CB1 and CB2 receptor immunoreactivity.
Whether the changes in the endogenous cannabinoid system in pancreatitis represent a beneficial response or cause the pathology has not been fully elucidated, and different results were obtained in animal species (rats and mice)—for example, CB1 and CB2 receptor activation reduces pathology associated with cerulein-induced pancreatitis in mice,120 whereas CB1 receptor activation in rat increased the severity of cerulein-induced pancreatitis and blockade of CB1 improved arterial pressure and survival rate.59 169 Dose differences may also contribute to these paradoxical findings, with low doses of cannabinoid agonists being protective and high doses aggravating pancreatitis. This is consistent with clinical reports of pancreatitis in heavy marijuana smokers.170 171
COLON, LIVER AND PANCREATIC CANCER
There are direct antineoplastic actions of cannabinoid agonists on several tumour cell lines in vitro and in animal models.172 173 The proposed mechanisms involve induction of apoptosis in tumour cells, antiproliferative actions, and antimetastatic effects through inhibition of angiogenesis and tumour cell migration.6 172 173 The antitumour actions may be mediated by activation of CB1, CB2 or a non-cannabinoid receptor-mediated mechanism.
Colon cancer
In human colorectal carcinoma cell lines, antiproliferative and apoptotic effects are largely mediated by CB1 receptor activation, though CB2 receptor activation cannot be definitely excluded.36 174 Activation of the CB1 receptor inhibited both RAS–MAPK/ERK and phosphatidylinositol 3-kinase (PI3K)–AKT survival signalling cascades, that are key cell survival pathways frequently dysregulated in colorectal tumours.170 Endocannabinoids may affect metastasis development since anandamide (via CB1 receptor activation) inhibits colon carcinoma cell migration.175 Oestrogens, which exert significant antiproliferative actions in colon cancer, upregulate CB1 receptor expression in human colon cancer cells.176
Anandamide may also exert selective antineoplastic actions through cannabinoid receptor-independent mechanisms including inhibition of tumour cell growth and induction of cell death in carcinoma cell lines that are high expressors of COX-2.177 This is relevant since COX-2 is highly expressed in the majority of colorectal cancers. The effect of anandamide was, in part, mediated by anandamide metabolites produced by COX-2.177
In vivo results support antineoplastic effects of cannabinoids in the digestive tract. Pharmacological inhibition of FAAH may be protective (via caspase-3 activation) against the preneoplastic lesions induced in the mouse colon by the genotoxic agent, azoxymethane.41 In an epidemiological review, marijuana smoke, which contains several of the carcinogens and co-carcinogens found in tobacco smoke, did not increase the risks of colorectal cancer in humans.178
Hepatocellular carcinoma
Cannabinoids modulate several signalling pathways involved in hepatic carcinogenesis, including activation of p38 mitogen-activated protein kinases and the PI3K–AKT survival pathways.179 In 64 primary hepatocellular carcinomas from patients, there was correlation between CB1/CB2 expression and two clinicopathological parameters (histopathological differentiation and portal vein invasion), with overexpression associated with improved prognosis.134
Cholangiocarcinoma
Endocannabinoids may affect cholangiocarcinoma growth. In cholangiocarcinoma cell lines which express TRPV1, CB1 and CB2 receptors,180 anandamide and 2-AG exert differential effects on cholangiocarcinoma cell growth via receptor-independent mechanisms. Anandamide caused antiproliferative and proapoptotic responses which were mediated by accumulation of ceramide and the recruitment of the Fas death receptor into the lipid rafts. In contrast, 2-AG exerted proliferative effects which were probably due to the disruption of the lipid raft structure and hence disturbance of the subsequent lipid raft-dependent cell signalling cascades.180
Pancreatic cancer
CB1 and CB2 receptors are overexpressed in areas of desmoplasia. CB2 immunoreactivity was evident in infiltrating mononuclear cells associated with pancreatic cancer.181 Cannabinoids (via CB2 receptor activation) induce apoptosis of pancreatic tumour cell lines in vitro and exert a remarkable growth-inhibitory effect in models of pancreatic cancer in vivo, in terms of both local invasion and distant metastases.182 There is a correlation between low cancer CB1 receptor or high FAAH/MAGL levels and survival181; this contrasts with improved prognosis observed in patients with hepatocellular carcinoma with overexpression of CB receptors174 (see above).
STRATEGIES TO ENHANCE CANNABINOID-BASED THERAPIES IN GASTROINTESTINAL AND LIVER DISEASES
Pharmacological considerations to avoid non-psychotropic effects associated with cannabinoid agonists
Central side effects associated with cannabinoids
The neuropsychiatric adverse effects of long-term use of Cannabis are well established, based mainly on observations from long-term marijuana smokers.183 These effects are largely mediated by activation of CB1, resulting in sedation, cognitive dysfunction and psychotropic effects. In clinical studies, therapeutic administration of Δ9-THC may provoke a sensation of a “high”, drowsiness, sedation and somnolence, euphoria, dizziness, dysphoria or depression, hallucination and paranoia.98 184 Hallucination (incidence: 6%) and paranoia (incidence: 5%) occurred exclusively with cannabinoids and were not observed in the control group.98
Development of cannabinoids drug devoid of central effects
Several approaches are being tested to avoid the unwanted side effects of cannabinoids. These include development of selective CB1 agonists that do not cross the blood–brain barrier; or selective targeting of CB2 receptors (since CB2 receptors are largely absent from the brain); or use of inhibitors of endocannabinoid inactivation (ie, FAAH inhibitors and inhibitors of endocannabinoid re-uptake) increasing tissue levels of anandamide to act on local CB1 receptors, without central effects. Potential applications of cannabinoid drugs in treatment of gastrointestinal, pancreatic and liver diseases are listed in table 3.
Cannabis-derived non-psychotropic constituents
Cannabis-derived non-psychotropic cannabinoids185 can avoid cannabinoid-associated central effects. The best studied among these compounds is cannabidiol (CBD), which is generally found in relatively high concentrations in the plant. CBD, unlike Δ9-THC, has very low affinity for both cannabinoid CB1 and CB2 receptors,186 although it has been proposed that it may modulate endocannabinoid function through its ability to inhibit the hydrolysis and the transport of anandamide and to act as a TRPV1 agonist.187 CBD has an extremely safe profile in humans188 189 and possesses pharmacological activities (eg, antioxidant, neuroprotective and anti-inflammatory actions)190 of potential interest in the gastrointestinal tract. However, effects of CBD in the gut are largely unexplored, with only a few studies published to date. CBD was effective in animal models of anticipatory nausea and vomiting (eg, conditioned gaping in rats and conditioned retching reactions elicited in the house musk shrew), suggesting potential in the treatment of chemotherapy-induced nausea.54 95 191–193 Ligresti and colleagues showed that CBD is a potent inhibitor of cell growth in neoplastic intestinal epithelial (Caco-2) cells.194 Finally, studies in rodents have shown that CBD did not modify motility in control animals,195 196 but it may inhibit the increased intestinal motility associated with experimental ileitis in mice.197
Genetic variation, gastrointestinal function and pharmacogenetic considerations
The functions of endocannabinoids and cannabinoid receptors are potentially altered by genetic variations, affecting the rate-limiting enzymes for synthesis or inactivation of CB receptors. These variations may potentially alter responses to cannabinoid modulation. For example, the natural C385A single nucleotide polymorphism (SNP) in the human FAAH gene produces a mutant enzyme which reduces the enzymatic activity of FAAH, which would result in higher anandamide levels, and stimulation of CB1 receptors, potentially impacting the efficacy of a CB receptor-modulating drug. The prevalence of the SNP is 16–25% in studies of Caucasians reported by the National Center for Biotechnology Information (NCBI) and in healthy controls and IBS patients in southeastern Minnesota.103 In a study of 482 patients with a range of functional gastrointestinal disorders and 252 healthy volunteers, there was a significant association of FAAH genotype with FGID phenotype and with specific individual phenotypes (specifically D-IBS and IBS with mixed bowel function), and with accelerated colonic transit in D-IBS. Pharmacogenetic studies in relation to gastrointestinal diseases are awaited.
There are as yet no reports of any clinically relevant effects of SNPs in the MGLL gene which encodes MAGL, the rate-limiting enzyme for metabolism of 2-AG, or in CNR1.
CONCLUSIONS
Experimental and human studies provide a substantial body of evidence that cannabinoid mechanisms are involved in several critically important functions in the gastrointestinal tract, liver and pancreas. These include motility, sensation, secretion, inflammation, tissue proliferation, carcinogenesis and fibrogenesis peripherally, and satiation, appetite and euphoria centrally. The potential therapeutic indications of cannabis have been hitherto based on myths, folklore, popular use and observations that cannabis is associated with “munchies” or gorging observed when people are high on marijuana. These diverse effects led to potential use of cannabinoid agents in a large variety of functional, inflammatory, or neoplastic diseases (table 3). We believe that application of modern pharmacological principles will expand the selective modulation of the cannabinoid system peripherally in humans. The approval in several countries of the CB1 antagonist, rimonabant, for the treatment of obesity and associated metabolic dysfunctions is the first selective cannabinoid modulator to have an impact on human disease. We anticipate a significant impact in other gastrointestinal and hepatic diseases associated with inflammation, fibrogenesis and neoplasia.
REFERENCES
Footnotes
Competing interests: None.